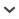To verify the common assumption that the provisioning of new nesting structures and/or artificial nest cups helps breeding populations of House Martins Delichon urbicum, we examined (1) the use of new self-standing nest site constructions (offering multiple artificial nest cups) erected to replace lost nesting sites or as additions, and (2) the extent to which House Martins breeding on buildings use artificial nests rather than self-built nest cups. We contrasted breeding performances, measured during two country-wide citizen science projects in The Netherlands, on two soil types, clay and sand, with clay being the expected preferred building material for nest cups over sand. The likelihood that artificial nest site constructions erected as replacements for lost nest sites were occupied was on average almost four times higher (occupancy rate 66%) than added structures (17%). Soil type had no significant effect on the occupancy rate. Confirming that clay is a better nest-building substrate than sand, self-built nest cups on sandy soils were more likely to collapse during a breeding season than nest cups built in areas with clay soils. Artificial nests had higher rates of occupation on sandy soils and in colonies where fewer self-built nest cups from previous seasons remained. On clay soils, self-built nests showed higher nesting success than artificial nests, with the reverse on sandy soils. The probability of second breeding attempts was higher in artificial nests than in self-built nests, but surprisingly this was only so on clay soils, with a strongly negative effect of first laying date. We conclude that, especially on sandy soils, the provisioning of artificial nest cups helps House Martins by enabling breeding and/or by improving breeding success. Where old breeding sites disappear, local populations can be maintained by providing ready-made structures instead. Although House Martin populations can thus be helped with nest sites and artificial nests, a comprehensive evaluation of limitations on the population warrants scrutiny of other factors, such as food, i.e. the supply of aerial insects.

Based on evidence from oral history (Blythe 1969, Evans 1971, Piersma 2016), monitoring since the mid-1960s in the UK (Woodward et al. 2020), along with a few older count series in the UK (in: Piersma 2016) and country-wide assessments in The Netherlands since 1967 (Philippona 1974, Leys 2002, van Dijk 2013), it has become clear that after the Second World War the breeding population of House Martins Delichon urbicum in western Europe made a nose-dive, with losses of 90% or more between 1950 and 1990. Since 1990, the Dutch population of House Martins has shown stability or modest recovery, with trends varying between regions (Boele et al. 2022). Human attempts to help House Martins usually involve the provisioning of nest site constructions (‘houses’) and/or the supplementation of artificial nest cups at existing nest sites. Both actions are based on the tacit assumption that suitable nest sites and good nest-building material are key factors that limit House Martin breeding populations.
The ideas underlying all these well-intended attempts to help breeding House Martins appear to have gone largely untested in explicit ways, although Kettel et al. (2021) examined the number of attempted broods and nest success in different types of nests across the UK. This is a missed opportunity, as the provisioning of constructions to accommodate small colonies of House Martins and the supplementation of artificial nests at (often previously used) breeding sites can both be considered ‘experiments’ to test the assumption that nest sites and suitable nest-building material are key factors currently limiting House Martin populations. If the availability of suitable nest sites is indeed limiting, upon the provisioning of artificial nest constructions (houses) and nest cups, we expect them to immediately be used. If the quality of nest-building material is limiting, on the basis of the finding that clay represents the preferred (and better) building material over sand (Papoulis et al. 2018), we expect that (1) the likelihood of House Martins occupying artificial nests is lower on clay soils than on sand and (2) indicators of breeding success, including the frequency of second breeding attempts, differ between clay and sand.
To test these predictions, we review the occupation of House Martin housing structures in The Netherlands in the period 2009–2021 (for a first analysis, see: van den Bremer et al. 2019). In addition, the results of a citizen-science survey of House Martin nests and nesting across The Netherlands in 2018–2020 (see van den Bremer et al. 2020) are analysed with respect to the timing of nesting, breeding success and the occurrence of second breeding attempts in self-built and artificial nest cups with local soil type (clay vs. sand) as the key environmental factor.
Figure 1.
Two examples of House Martin houses, consisting of a ‘roof overhang’ where several artificial nests are placed, mounted on a pole. The construction of the roof overhang can differ between houses. (A) A more ‘traditional’ house that is most commonly used (photo Martine Dubois) and (B) a house using masonry stones (photo Johan Drop).

METHODS
Occupation of House Martin houses
To help replace lost nest sites, or simply to help House Martins, people have started providing man-made nesting structures (‘houses’). A House Martin house consists of a ‘roof overhang’ where artificial nests are placed, mounted on a pole (Figure 1). Based on the volunteer efforts as described by de Jong & van Berkel (2013), information on location, date of placement and annual occupation was available for the years 2009–2021 for 264 House Martin houses (see Figure 2). For a subset of the houses, note was made of the motivation for placement of the House Martin houses, i.e. whether they were built to compensate for the loss of original nesting locations, or whether they were simply an addition. In some cases, House Martin sound recordings were used to attract birds to the structures, but this was not noted systematically for the vast majority of locations. Selecting the 248 structures on clay or sandy soils (and thus omitting 16 houses on peaty soils from the analysis, see below), soil type was based on geodata ( www.pdok.nl).
National House Martin nest study
To quantify breeding performance of the Dutch breeding population of House Martin, a national House Martin nest study was organised in the years 2018, 2019 and 2020. During April–September volunteer observers across The Netherlands were asked to make weekly observations of nest activity using standardized observations based on Piersma (2013) and simplified by van den Bremer et al. (2020; and see Kettel et al. 2021). Any object or building with one or more nests that were clearly visible from the ground could be a study location. During the first visit to the colony in mid-April, the initial situation of the study site was described. The exact geographic location of the study location, the type of object hosting the House Martins, the number of artificial nests (Figure 3) and the presence of remaining complete and incomplete old self-built nests of the previous breeding season were registered. From late April onwards the study location was visited weekly and the ‘breeding stage’ of all nests or a sample of nests was noted during 1-hour observation sessions, using a uniform list of nest codes (Table S1).
Figure 2.
Distribution of artificial nest site constructions (House Martin ‘houses’; squares) in 2009–2021 and locations of colonies (circles; colours represent number of years studied, size represents number of nests within each colony) where nests were scrutinized during the House Martin nest study in 2018–2020 across The Netherlands, in relation to soil type (sand or clay).


On this basis we determined the total number of occupied nests, the proportion of successful nests and the proportion of nests in which a second breeding attempt was started (not necessarily by the same two partners, TP pers. obs. based on ringed individuals, hence we do not call it ‘second brood’), after the first was either successful or failed. The date of the onset of laying (lay date) was estimated on the basis of observations in the egg and young phase, where we assumed that a full breeding cycle (from laying the first egg to fledging young) lasts 44 days (egg laying 5 days, brooding 15 days, young 24 days; Cramp 1988). With visual observations of the nests available only, it was not possible to assess the number of hatched or fledged young (Piersma 2013). Based on the efforts of 115 volunteers, in 2018 we obtained observations of 1037 breeding attempts at 90 study locations, in 2019 observations on 621 breeding attempts at 46 study locations and in 2020 observations on 571 breeding attempts at 41 study locations. The study locations showed a good spread across The Netherlands (Figure 2). Soil type (sand or clay) was determined for each location using geodata ( www.pdok.nl); note that the data on 196 nests collected on peaty soils were left out of the analysis. There was minimal overlap in locations between the National House Martin nest study and the study on the occupation of House Martin houses.
Statistics
To determine which factors had an effect on the occupation of House Martin houses (with a house being the unit of measurement and analysis), we fitted a Generalized Linear Model (GLM) with binomial distribution, using occupation (‘yes’ or ‘no’) as response variable, and soil type (sand or clay) and the number of years since placement of the House Martin house as explanatory variables, including their interaction. To be clear, the number of years since placement equals the number of years that the house has been present at each location. A House Martin house was considered ‘occupied’ when during at least one of the years since placement at least one nest on the house was occupied. In a second, separate model, we added motivation (compensation or ‘extra’) as a third explanatory variable, as this motivation was available for only a part of the data (n = 175 houses). The use of sound was available for less than 10% of the data and could not be included in the analysis.
With individual nests in the National House Martin nest study as the units of measurement, we calculated nest success (i.e. the proportion of nests in which at least one young fledged), for first and second breeding attempts separately, with the binomial loglinear regression model that was described by Aebischer (1999). This method is also referred to as Mayfield Logistic Regression (Hazler 2004) and is an extension of the Mayfield method (Mayfield 1975). Mayfield's method accounts for number of ‘exposure days’, i.e. the number of days during which a nest is under observation, from the time it is found until it fails, fledges, or is censored. For a group of nests, the daily failure rate r is F/E, where F is total number of nest failures and E is number of exposure days summed over all nests. Daily survival rate S is then 1 – r, and probability of a nest surviving for a nesting cycle of d days is Sd (here 44 days). Mayfield Logistic Regression extends this traditional Mayfield estimator by (1) using the framework of Generalized Linear Modelling for fitting Mayfield models (GLM with logistic link function and binomial error term), where the unit of analysis is the nest and the response variable is the number of days of observation during which that nest is successful, and (2) incorporating contextual variables for each nest (Aebischer 1999, Hazler 2004).
The quality of sand or clay as a building material for nests was investigated by examining the effect of soil type on (1) the frequency of self-built nests collapsing within the breeding season, (2) the proportion of successful nests, i.e. nest success, assessed separately for first and second breeding attempts, and (3) the probability of a second breeding attempt in a nest. To assess whether soil type has an effect on the number of self-built nests collapsing, we fitted a GLM with binomial distribution. Next to soil type (sand or clay), clutch number, year and their interactions were also fitted to the model. To examine whether soil type had an effect on nest success, we modelled daily nest survival (response variable) in relation to nest type (self-built or artificial), soil type (clay or sand) and whether it was the first or second nesting attempt of the season, including all interactions. Year was added as a factor. In all analyses non-significant (interaction) terms remained in the model when testing and estimating the effects of the single terms. In the same way, a GLM was run to assess the effect of soil type on the probability of a second nesting attempt, whilst lay date of the first clutch was also included as an explanatory variable.
The statistical effect of soil type on the supply of old self-built nests in April/May was tested using a GLM with Poisson distribution, with a nesting location (or colony) as the unit of analysis. Next the effect of the availability (i.e. the number) of self-built nests in April/May remaining from the previous breeding season on the proportion of occupied artificial nests was tested using a GLM with a binomial distribution, using year and soil type as additional variables. The number of artificial nests in April was used as the binomial totals in the denominator. All statistical analyses were carried out in GenStat v. 18 (VSN International 2015).
RESULTS
The probability that a House Martin house became occupied was not significantly higher on sandy soils (mean ± SE: 27% ± 4.1) than on clay (21% ± 4.0; Table 1). There was some indication that the likelihood of occupation increased with the number of years after placement (estimate ± SE: 0.11 ± 0.07: from 12% ± 5.8 in first year after placement to 33% ± 10.3 in year 13 on clay; from 25% ± 8.2 in first year after placement to 30% ± 8.8 in year 13 on sand), but this effect was not statistically significant (Table 1). In the second analysis with a reduced dataset including motivation, the occupation rate of houses placed as a compensation measure (because the original nesting location had disappeared or was made inaccessible) was significantly higher than of houses placed as additions only (on average 66% ± 8.4 vs. 17% ± 3.1; Figure 4, Table 1). Note that in this latter analysis soil type and number of years after placement had no effects on occupation rate either (Table 1).
The proportion of individual nests that collapsed during the breeding season was significantly higher on sandy than on clay soils (t9 = 4.15, P = 0.002; Figure 5). The proportion of nests that collapsed did not statistically differ between first and second breeding attempts (t9 = 1.45, P = 0.18), and there were no significant effects of the interactions between covariates. Similarly, the proportion of artificial nests that was occupied tended to be larger on sandy soils than on clay soils (with significance not reaching the 5% confidence level in a two-sided test; t79 = 1.79, P = 0.08; Figure 6A). The availability of old self-built nests at the start of the breeding season was on average higher on clay soils than on sandy soils (t79 = –7.41, P = 0.001), with fewer artificial nests becoming occupied when more self-built nests from previous breeding seasons remained (t79 = –2.22, P < 0.001; Figure 6B).
Figure 4.
Mean observed occupancy rate of House Martin houses ± SE per type of motivation for placement (compensation, n = 33, or extra, n = 142, see text) based on the model output as described in Table 1.

Figure 5.
Proportion of natural nests that collapsed per soil type and clutch number (± SE). Sample size (number of nests collapsed / total number of nests) per soil type and clutch number is given.

Figure 6.
(A) Mean observed occupancy rates (± SE) of artificial nests in July on sandy and clay soils and (B) occupancy rate of artificial nests in July in relation to the availability of old self-built nests at the start of the breeding season in April, given for sandy soils and clay soils.

Table 1.
Statistics of a generalized linear multivariate model estimating the effect of soil type and the number of years after placement on the occupancy of House Martin houses (Model 1, n = 248 houses, df = 244), including their interaction, with and without ‘motivation’ as explanatory variable (Model 2, n = 175 houses, df = 170).

Nest success was higher in self-built nests than in artificial nests (t2217 = –4.33, P < 0.001), but only on clay soils (interaction significant; t2217 = 6.10, P < 0.001; Figure 7 lower panel). As a result, overall nest success of first clutches was highest in self-built nests on clay soils (88.4%), and lowest in self-built nests on sandy soils (77.1%). Across soil and nest types, nest success was higher for first than for second breeding attempts (t2225 = –4.25, P < 0.001).
The proportion of nests hosting a second brood was lower for self-built nests than for artificial nests (t1582 = 2.55, P = 0.011), but only on clay soils (interaction significant; t1582 = –2.43, P = 0.015; Figure 7 upper panel). The best predictor of the proportion of second breeding attempts, however, was the date of egg-laying: the earlier the first breeding attempt, the higher the likelihood of a second one (estimate = –0.047, t1210 = –10.4, P < 0.001; Figure 8). Laying date of the first clutch was strongly determined by nest type (t1208 = –4.25, P < 0.001) and the interaction between nest type and soil type (t1208 = 3.43, P < 0.001): in artificial nests House Martins started egg laying on average 6.8 days earlier than in self-built nests, but only on clay soils (day ± SE: 155.9 ± 1.44 vs. 162.7 ± 0.695).
Figure 7.
Probability of second broods per soil type (clay, sand) and nest type (self-built, artificial; ± SE). Percentage of successful nests (at least one fledged young; Mayfield, including 95% confidence limits) for first and second clutches per soil type (clay, sand) and nest type (self-built and artificial). Numbers of nests per category are given above the bars.


DISCUSSION
The findings that newly erected nest site structures had low occupancy on both clay and sand, and that occupancy of a House Martin house did not significantly increase over years, do not suggest a strong lack of nest sites or nest-building materials for House Martins breeding in The Netherlands. However, the finding that new martin houses replacing lost nest site structures had an almost four times higher occupancy rate than added ones, may count as evidence for faithfulness to a breeding site. The replaced breeding structure may reflect the quality of a site in terms of food and safety.
The results on individual nests are consistent with the idea that, compared to sand, clay soils provide House Martins with the better nest-building material, resulting in fewer intra-seasonal nest collapses, and more self-built nests remaining from previous years. This finding also tallies well with nest-building material selection in swallow species (Papoulis et al. 2018) and with our finding that on clay soils House Martins prefer self-built nests and have higher nest success during the first nesting attempt. It is also consistent with the observation, actually in one of the colonies in the survey (on clay soil), that only when the availability, upon spring arrival, of the preferred old clay-nests was low, House Martins occupied artificial nests (Piersma 2013). The finding that only on clay soils the likelihood of second broods was higher in artificial than in self-built nests is at odds with the idea that second broods indicate occupation of that nest by early-arriving and older birds (Piersma 2013). Although soil type was not taken into account in an analysis for the UK (Kettel et al. 2021), here House Martins were also more likely to attempt two broods and to succeed in nesting attempts in artificial nests compared with natural nests.
On sandy soils, where appropriate nest-building material is harder to find than on clay (see Dijkstra 2013), House Martins tend to accept artificial nest sites more readily as an alternative than on clay. In fact, the artificial alternative appears to be good, as on sandy soils nest survival was higher in artificial nests than in self-built nests. Of course, this still does not mean that populations on sandy soils are necessarily limited by suitable nesting material, they may just take it easier if such alternatives are provided and do well as a result. What holds for sandy soils may also hold for peat soils (for which our samples sizes were too small for analysis).
This study adds to the ones of Willi et al. (2011) and Kettel et al. (2021) in confirming the idea that the addition of nest site structures and/or artificial nests helps House Martins to make successful breeding attempts, especially on sandy soils. That only a low percentage of newly added artificial houses becomes occupied is not consistent with the idea that lack of nest sites and/or nest-building material currently limit the northwest European House Martin population, let alone explain their strong historical decline. On the other hand, since particularly on sandy soils the majority of artificial nest cups becomes occupied and their nest success is even higher than of natural nests, it is conceivable that the provision of artificial nest cups contributed to halt the steep population decline from the 1990s onwards. Full explorations of the factors limiting House Martin populations should especially include the abundance of aerial arthropod food (Newton 2004, Nebel et al. 2010, Hallmann et al. 2014) and their availability, e.g. in relation to weather (Finch et al. 2023), and monitor changes in demographic parameters that were not assessed in this study (notably the number of fledged young). Finally, by way of practical advice, we now know that artificial nests will help House Martins on some soils (i.e. sand, and perhaps peat) that fail to offer the best nest-building materials, and that new housing structures will help where traditional nest sites have been removed.
ACKNOWLEDGEMENTS
We are very grateful to the hundreds of volunteers who participated in the National House Martin nest study and kept track of the occupation of House Martin houses. This work would not have been possible without their impressive effort of weekly nest observations for many years. We thank Albert de Jong for his support in the coordination of the fieldwork, Jeroen Nienhuis for his help with the data analysis and Lara Marx for providing the distribution map. We thank Hans Schekkerman for his advice in the statistical analysis. We are grateful for Wilfried de Jong (†) and Will van Berkel who monitored the occupation of House Martin houses for many years, which was later taken over by Hans Willemsen in cooperation with Birdlife Netherlands. This study has been made possible by the financial support of Birdlife Netherlands and the Prins Bernard Cultuur Fonds. We thank Rob Bijlsma and anonymous reviewers for comments on earlier versions of the manuscript.
REFERENCES
Appendices
SAMENVATTING
Mensen proberen lokale populaties van Huiszwaluwen Delichon urbicum doorgaans te helpen door de vogels additionele nestgelegenheid aan te bieden door het plaatsen van een huiszwaluwtil of het bijplaatsen van kunstnesten. Een zwaluwtil is een soort dakoverstek op een vrij hoge paal waartegen kunstnesten zijn aangebracht. Beide maatregelen zijn gebaseerd op de veronderstelling dat geschikte nestplaatsen en goed nestbouwmateriaal sleutelfactoren zijn die broedpopulaties van Huiszwaluwen beperken. Wij onderzochten (1) de bezetting van huiszwaluwtillen die geplaatst waren ter compensatie van verdwenen nestgelegenheid of als extra nestgelegenheid en (2) de mate waarin Huiszwaluwen die tegen gebouwen broeden, kunstnesten in plaats van zelfgebouwde nesten gebruiken. Op grond van georganiseerde en gestandaardiseerde waarnemingen tijdens twee landelijke burgerwetenschapsprojecten in Nederland (2009–2021, 2018–2020) hebben we de broedprestaties vergeleken in gebieden met twee verschillende grondsoorten (klei of zand), waarbij klei het verwachte bouwmateriaal is dat de voorkeur heeft voor nestbouw boven zand. De kans dat een huiszwaluwtil geplaatst als compensatiemaatregel bezet raakte, was gemiddeld bijna vier keer zo groot (66%) als bij tillen die als extra nestgelegenheid waren geplaatst (17%). De grondsoort had geen significant effect op de bezettingskans van een huiszwaluwtil. Zelfgebouwde nesten op zand hadden een grotere kans om in te storten gedurende het broedseizoen dan zelfgebouwde nesten in gebieden met kleigrond, wat bevestigt dat klei beter nestbouwmateriaal is dan zand. Kunstnesten hadden een hogere bezettingsgraad op zandgrond en in kolonies waar er minder zelfgebouwde nesten van het vorige broedseizoen aanwezig waren. Op kleigrond hadden zelfgebouwde nesten een groter nestsucces dan kunstnesten, terwijl op zandgrond het omgekeerde het geval was. De kans op een tweede broedpoging was groter in kunstnesten dan in zelfgebouwde nesten, maar verrassend genoeg alleen op kleigrond, met een sterk negatief effect van de eerste legdatum. We concluderen dat, vooral op zandgrond, het aanbieden van kunstnesten Huiszwaluwen kan helpen aan nestgelegenheid en/of het verbeteren van het broedsucces. Waar oude broedplaatsen verdwijnen, kunnen lokale populaties geholpen worden door het plaatsen van huiszwaluwtillen. Hoewel Huiszwaluwen dus kunnen worden geholpen met kunstmatige nestplaatsen en nesten, is een uitgebreidere analyse van de sturende factoren van de populaties van belang, waarbij ook andere factoren, waaronder voedselaanbod (vliegende insecten), worden onderzocht.