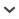Like many other terns, European Sandwich Terns Thalasseus sandvicensis exhibit a period of post-breeding dispersal before migrating to their wintering grounds in Africa. To understand the significance of Danish waters for these movements, we analysed changes in the abundance and distribution of Sandwich Terns throughout Denmark from late June to October using our own count data as well as data from the online bird portal DOFbasen. To understand the geomorphology of, and threats to important roosting sites, we assessed their habitat characteristics and the level of (human) disturbances using data recorded in the field. We used 2422 resightings of individually marked juvenile Sandwich Terns to assess the use of Danish waters by terns of different geographical origin. The results showed that the temporal use of roosting sites differed between the western and eastern part of Denmark, with maximum numbers of Sandwich Terns occurring earlier in the west. Important roosting sites were characterised by sandy shallow coastlines, which reflect suitable habitats for their key food species. It seems most likely that phenological patterns in Sandwich Terns reflect the phenology and relative abundance of their food items. Results from the resighting data suggested that the proximity of breeding colonies to the different post-breeding areas in Denmark influenced the choice of post-breeding area as well as the phenological patterns. Large roosting numbers in combination with resightings of Sandwich Terns dispersing from various European countries highlight the international importance of Danish waters during the period of post-breeding dispersal. Observed average disturbance rates exceeding one disturbance per hour underlines the need for protected roosting locations on beaches with high human activity.
Many bird species migrate to exploit seasonal resource availability in different parts of the world at different times in their annual cycle and have developed different strategies to accomplish their migratory journeys (Alerstam et al. 2003, Alerstam 2011). Migratory schedules may differ between species, and within these schedules, post-breeding dispersal emerges as a potential strategy to enhance fitness. Post-breeding dispersal describes movements after breeding but prior to the actual autumn migration to the wintering grounds. The dispersal may have several functions, including locating and profiting from rich feeding areas when fattening up before migration and exploring resource availability in alternative future breeding areas. The phenomenon is observed in several bird species and can play an important role in shaping distributions and population dynamics (Bowler & Benton 2004, Newton 2008). However, the specific triggers for post-breeding dispersal vary among species and different ecological and physiological factors can contribute to this behaviour. While some species use the opportunity to explore more favourable food resources or suitable habitats and potential breeding sites before migration, other species do not engage in post breeding dispersal, e.g. due to limited time or energy resources.
As a migratory seabird, Sandwich Terns Thalasseus sandvicensis are known for their post-breeding dispersal (Møller 1981, Cabot & Nisbet 2013, Fijn et al. 2014), whereby juvenile birds and their attentive parents depart the vicinity of the natal colony to feed and remain at sites elsewhere, prior to the initiation of active migration. Post-breeding dispersal to such prey-rich areas likely has multiple potential benefits for Sandwich Terns. For example, it could permit (1) learning of complex feeding methods by juveniles (Dunn 1971, Cabot & Nisbet 2013). Like many other terns such as Elegant Terns Thalasseus elegans, Royal Terns Thalasseus maximus and Roseate Terns Sterna dougallii, Sandwich Terns provide prolonged parental care and feed their offspring throughout post-breeding. This is thought necessary due to their complex feeding technique, ‘plunge-diving’ to feed on schooling fish, which can take months for the juveniles to practise and master (Ashmole & Tover 1968, Shealer & Kress 1994, Cabot & Nisbet 2013). (2) Flight feather moult and replacing worn feathers enhances adult flight performance prior to autumn migration (Ward 2000, Meissner & Krupa 2007, Jenni & Winkler 2020). (3) The long flight to African wintering grounds requires energy reserve acquisition (Ward 2000, Meissner & Krupa 2007). Finally, (4) prospecting in potential future breeding areas provides important knowledge not only for failed breeders but also for naïve juveniles and successful breeders (Cabot & Nisbet 2013, Fijn et al. 2014). Sandwich Tern colonies can be highly unstable, and birds often shift to new locations due to predation, food shortage, habitat destruction or relocation of Black-headed Gull Chroicocephalus ridibundus colonies, with which Sandwich Terns usually associate (Herrmann et al. 2008, Cabot & Nisbet 2013, Shealer et al. 2020). The capacity to make such shifts based upon previous experiences (e.g. failed breeding) as well as current experiences (e.g. disturbances before or during breeding) crucially depends upon the collective knowledge gained from exploring potential breeding sites during post-breeding dispersal.
Mature Sandwich Terns from colonies in northwestern Europe usually return from their wintering grounds around late March and start breeding by the end of April. Depending on the success of the brood, the period of post-breeding dispersal starts around late June and can last well into September, with a few individuals remaining even into October at the post-breeding sites in the North and Baltic Seas (Møller 1981, Cabot & Nisbet 2013). However, by October, most birds have arrived in their wintering quarters in West Africa and in Southwest Africa by November. The north-western European Sandwich Terns from Denmark, Germany, The Netherlands, Great Britain, Poland and Sweden, all derive from the same meta-population, showing high levels of dispersal between colonies and visiting different countries during post-breeding, including movements to the north of their breeding or natal colonies (Møller 1981, Cabot & Nisbet 2013, Fijn et al. 2014).
Globally, Sandwich Terns are not threatened, being classified as Least Concern (LC) on the IUCN Red List (BirdLife International 2021). However, despite overall increases in the Atlantic population since the early 2000's (current population estimate: 170,000–200,000 individuals; Wetlands International 2021), breeding numbers have recently declined, due to avian influenza outbreaks that caused extensive mortality among adult breeders in several of the largest breeding colonies in the North Sea region (Rijks et al. 2022, Knief et al. 2024). Furthermore, the E.U. Birds Directive (2009/ 147/EC) lists Sandwich Terns on Annex 1 as specially protected, because of their sensitivity to environmental changes, disturbances, predation and changing food availability (European Union n.d.). Sandwich Tern conservation efforts have concentrated on protection of suitable breeding sites (Baltic Marine Environment Protection Commission 2013). However, effective conservation measures during post-breeding dispersal might also influence breeding population size in subsequent years. Foraging and roosting Sandwich Terns select sandy beaches with adjacent shallow waters (Cabot & Nisbet 2013, Shealer et al. 2020). Although less threatened than some of their breeding locations, such habitats are highly attractive to humans especially during summer months, which coincides precisely with post-breeding dispersal, causing intense anthropogenic disturbance to beach-roosting terns.
The coast of Denmark is characterised by stretches of extensive sandy shores and sandbanks associated with shallow inshore waters, conditions offering highly suitable roosting and foraging places for Sandwich Terns (Cabot & Nisbet 2013). Ringing records have shown that many north-western European Sandwich Terns disperse to different parts of Denmark, probably to profit from its rich feeding grounds (Haupt 2011, Cabot & Nisbet 2013, Fijn et al. 2014). Sandwich Terns mainly feed on sandeel Ammodytes sp., Sprat Sprattus sprattus and herring Clupea sp., which use Danish waters as spawning and nursery areas during summer (Daan et al. 1990, Cabot & Nisbet 2013). However, little is known about the significance of different areas across Denmark for Sandwich Terns dispersing during the post-breeding period.
Here, we provide information about the phenology, geographical origin and use of Danish waters by Sandwich Terns throughout the period of post-breeding dispersal. We analysed the phenology at roosting sites that were identified as being important and we investigated (1) site-based maximum numbers, (2) variation in abundance between years and (3) differences in phenology patterns between sites and regions. Furthermore, we looked at specific roosting sites in order to understand their geomorphology and identify characteristics that were important to Sandwich Terns. Because some Danish roosting sites attract internationally important numbers of Sandwich Terns (Bregnballe et al. 2016), it is necessary to assess potential threats to promote the effective protection of valuable roosting sites, especially regarding levels of human disturbance at roosting sites during the post-breeding period.
Juvenile Sandwich Tern wearing a white colour ring. Juvenile Sandwich Terns can be recognized by the black pattern on their back feathers and the lack of yellow on the beak (Fanø, 20 July 2020).

Juvenile Sandwich Tern during post-breeding season wearing a vertical metal ring, roosting in a temporary pond on the beach (Hirtshals, 21 July 2020).

Finally, we investigated resighting data from individually marked Sandwich Terns recorded in Denmark. Møller (1981) provided initial information about dispersal to Denmark in relation to their European sites of origin, based only on ring recoveries of dead birds. Here, we undertake more detailed analyses, using resighting data from living marked birds to assess the importance of Danish waters for birds of differing geographical origins. We specifically tested the hypothesis that Sandwich Terns from different origins may associate with different areas in Denmark, e.g. that birds from the Baltic populations are more often seen in eastern Danish waters, closer to their breeding colonies. In this way, we assessed spatial and temporal dispersal patterns for different sub-populations of Sandwich Terns.
METHODS
Study area
This study took place at five pre-selected Danish coastal areas concentrated on nine sites, which we identified as being important roosting sites for Sandwich Terns, all of which supported consistently high numbers of roosting birds in relation to the general distribution in Denmark between 2000 and 2020 (Figure 1A, B). Two sites were in southwest Jutland (Fanø and Blåvand), one further north along the west coast (Agger Tange), two on the north coast (the coast stretches between Hirtshals and Skiveren, hereafter referred to as ‘Hirtshals Coast’, and Skagen) and three along the east coast (Nordmandshage, the north coast of Djursland from Udbyhøj to Gjerrild, and Ebeltoft). Numbers reported from Gedser in southeast Denmark were also included, where most birds roost on Rødsand, a sandy island (difficult to access and not covered by counts during summer/autumn) near Gedser; counts from Gedser provide a reflective impression of tern phenology in this area. Further characteristics of the individual study sites are described in the Results section.
Roosting sites
To identify characteristics of relevant roosting sites for Sandwich Terns and to understand what structures the terns were using for roosting, we collected geomorphological data from all nine study sites. We recorded habitat type and the specific roosting spots of the terns at the sites we visited. Information from Gedser/ Rødsand was provided by local birdwatchers.
We determined levels of human disturbance to roosting Sandwich Terns on beaches at five of the nine study sites which we had identified as popular Sandwich Tern roost sites as well as tourist hotspots (Fanø (SW), Blåvand (SW), Hirtshals (N), Djursland (E) and Ebeltoft (E)) during 77 observation hours on 19 days in July and August 2020. At Skagen (N), Sandwich Terns usually left the roosting site before the start of human activity, so that incidences of direct disturbance were difficult to capture. Disturbances were defined as events that provoked the terns to take off from their roosting point. During such events, we recorded the source of disturbance, the frequency of each type during an observation day and behavioural responses of the terns, e.g. whether they flew round and returned or left their previous roosting place. As data relating to disturbance on Sandwich Terns in Denmark had not been collected before, we could only include 2020 data that we first recorded in this study. We are aware there may be year-specific biases associated as a result. Although human activity continued to be influenced by the COVID-19 outbreak during 2020, in that summer, restrictions on human movements in Denmark were relaxed because of relatively low infection rates, although there were fewer summer tourists from other countries than normal. As such, especially for Danes, visits to nature and beaches were particularly popular, offering the opportunity to undertake studies of disturbance rates.
Figure 1.
Locations of Sandwich Terns that were relevant for this study. (A) Locations of nine important roosting sites for Sandwich Terns in Denmark, which were studied during this project. Study sites are grouped into five coastal areas, namely Southwest, West, North, East and Southeast. The north coast of Djursland and Hirtshals Coast were both spread over stretches of coast. (B) Overall distribution of roosting locations of Sandwich Terns around the Danish coast from 2000–2020, showing the highest number of Sandwich Terns recorded at each site. Dots with different colours and sizes indicate different maximum numbers of Sandwich Terns observed. Locations with high roosting numbers represent the nine study sites. (C) Locations of resightings of individually marked Sandwich Terns in Denmark grouped into the three coastal areas used in the present analysis of resightings of juvenile birds. (D) Locations of breeding colonies of Sandwich Terns grouped into ringing areas. Only breeding colonies from which ringed individuals had been resighted as juveniles in Denmark are presented.

Phenology and overall distribution of post-breeding birds
Because the first successful breeding pairs usually leave the Danish colonies by the end of June (K.T. Pedersen & J. Sterup pers. comm.) and a few birds remain in Denmark into October (Møller 1981), we used Sandwich Tern data collected between June and October to access their post-breeding dispersal. However, as autumn migration for most Sandwich Terns begins around September (Møller 1981, Cabot & Nisbet 2013, Shealer et al. 2020), we defined the end of June until the end of September as the main part of the post-breeding period.
The online bird observation portal DOFbasen ( www.dofbasen.dk), organized by the Danish ornithological association (Dansk Ornitologisk Forening), contains a large dataset on Sandwich Terns counted at coastal sites throughout Denmark by volunteer observers over many years. All freely available Sandwich Tern numbers, including those counted by the authors, can be found in this database. Most locations in DOFbasen contain data since the early 2000s, although for a few locations like Blåvand Bird Observatory (SW), data are available back to the 1960s. To be able to describe phenological patterns in different parts of Denmark, we summarised the DOFbasen data by identifying maximum numbers of roosting Sandwich Terns within 10-day-intervals (decades).
We divided each of the five months of the post-breeding period into three 10-day-periods (the last decade covering 10 or 11 days dependent on month), assigned each Sandwich Tern report to the corresponding decade and selected the highest roosting number recorded per decade. Occasionally night roost counts carried out at sunrise or late evening were excluded to avoid duplicate counting on the same day at day-roosts, because birds recorded at different locations during daytime may gather at communal roosts for the night, that were not used during the day (Sandwich Terns do not necessarily use the same roosting site during the day and night). We included records of Sandwich Terns associated with roosting or foraging behaviour, as well as records of flying individuals and records with unspecified behaviour, because we judged most birds in these categories were foraging associated with these roosts or roosting (an assumption which was sometimes confirmed by descriptions given in the observer-comments). These two behaviours accounted for a maximum of 10% of the data for each of the study sites, usually less. Only Gedser (SE) had a higher amount (c. 38%), as expected because most Sandwich Terns pass Gedser on foraging flights to and from nearby Rødsand, an assumption confirmed by associated comments.
We compared the post-breeding phenology between the nine study sites. Most of the individual study sites, except Blåvand (SW) and Skagen (N), consisted of several locations in DOFbasen. For the days where multiple counters had counted Sandwich Terns at the same location, we used the maximum of the day. For the areas where there were records from several neighbouring locations, we carefully evaluated whether it would be reasonable to the add numbers from the different locations to a sum for the entire study area. When doing this, we inspected the timing of the counts at the different locations in order to minimise the risk that the same birds would be included twice because of movements among sites within the day. In the next step, we extracted the maximum number per decade for each study site. To avoid inclusion of years with a poor coverage, we excluded years from which fewer than 60% of the decades of the main part of the post-breeding period (end of June until the end of September) contained count data. Depending on the study site, between 12 and 42 years of data were available (Table S1). Based on this selection, we obtained a general phenology pattern for each study site by calculating the average number of Sandwich Terns over all usable years for each decade. However, almost no information about zero-counts was available and it was not apparent whether missing data meant that the site was not counted or that there were no terns. Therefore, we only included the maximum number of roosting Sandwich Terns at a site per year in the comparative analysis.
Resighting data
Due to extensive ringing of nestlings in Denmark and neighbouring countries, it was possible to identify the geographical origin of many of dispersing juvenile individuals occurring in Denmark during post-breeding to assess spatiotemporal dispersal patterns of different north-western European colonies. As part of the study, we conducted several visits to Fanø (SW), Hirtshals (N), Ebeltoft (E) and North Djursland (E) to search for individually marked birds. We conducted searches for marked individuals annually using a telescope or camera (Swarovski ATX 95, Nikon P900) on Fanø (SW; 2016–2020), at Ebeltoft (E; 2018–2020) and at North Djursland (E; 2020) and along the coast of Hirtshals (N; 2020). The largest effort invested in the search of ringed individuals was made on Fanø (SW; almost daily searches), whereas searches for ringed birds at other sites were made approximately once per week.
In addition to our own resightings, Copenhagen Bird Ringing Centre as well as private databases provided further data and information (e.g. life history) of ringed Sandwich Terns that had been resighted, recaptured or found dead in Denmark. Infrequent resightings from voluntary observers were available for various sites since 2011. For the analyses, we only included data on birds that had been resighted alive during the 10 years from 2011 up to and including 2020. Adult Sandwich Terns do not necessarily breed in the area where they hatched and can shift colonies between years (Shealer et al. 2020), so to be certain about the colony location to which an individual belonged, we only included birds ringed as pulli and resighted as juveniles. Hence, we only examined resightings of the first year of each juvenile, which reflects movements of families given the prolonged parental care (Cabot & Nisbet 2013). Thus, in the following we use resighting data in order to examine the post-breeding dispersal of families. Because there were scarcely any birds that were both resighted breeding in a colony and then resighted in Denmark in the same year, we decided not to include resighting data of adult Sandwich Tern. For every resighted juvenile, we organised information about ringing date and location, age at ringing, as well as resighting location and age at resighting. This dataset was supplemented by records stored by the Dutch Sandwich Tern ringing group (Vogeltrekstation Arnhem, Heteren) and a ringing group at the northwest coast of Germany (Vogelwarte Helgoland, Wilhelmshaven), all providing information about individuals that had been recorded in Denmark. Due to a higher resighting probability based on our own experience, we only included data from individuals either marked with colour rings or special vertical rings (tall steel rings with six vertical letters/numbers). Therefore, we did not evaluate resightings of Sandwich Terns marked with smaller traditional steel rings with horizontal digits, resulting in the exclusion of 86 resightings from our analysis, amongst them all the resightings of terns from colonies in Sweden (n = 11).
As there were only a few resightings from the western or south-eastern coastal area of Denmark, we divided the resighting locations into three major coastal areas for further resighting analysis: Southwest, North and East Denmark (Figure 1C). To determine the origin of the resighted birds, we looked at the location of their different hatching colonies. However, we grouped together those colonies in a country that were in close proximity to each other and/or from which only a few birds were resighted. Therefore, we grouped ringing locations into the following 10 areas: the southeast coast of Denmark (three colonies close to Funen, plus one nearby colony on Zealand with few resightings), the west coast of Denmark (one colony), the island of Hirsholm (one colony; Northern Kattegat, Denmark), Hallig Norderoog (one colony; Germany), the island of Baltrum (one colony; Germany), the northeast coast of Germany (one colony), the northwest coast of The Netherlands (10 colonies in close proximity in the Wadden Sea, plus one nearby colony in the North Sea with few resightings), the Delta region in The Netherlands (three colonies in close proximity), Great Britain (three colonies with one resighting each) and Poland (one colony; Figure 1D). To look for spatiotemporal patterns of different breeding populations in the post-breeding areas, we selected ringing areas from where at least 10 different individuals had been resighted in at least one of the major post-breeding areas (Southwest, North and East Denmark; see Figure 1C). Again, the post-breeding period was divided into decades.
Data analysis
All statistical analyses were performed in R (R Core Team 2021).
To test for differences in timing of maximum numbers among study sites, we fitted a linear mixed model by using maximum likelihood in the ‘lme4’ package in R (Bates et al. 2015) with decade (temporal parameter across years) as dependent variable, site (spatial parameter) as fixed and year as random factor. To compare variation between years, we only included records from 2008 to 2020, as data for almost all sites were available for those years (for at least seven out of nine sites). By using the ‘ranova’ function in the ‘lmerTest’ package (Kuznetsova et al. 2017), we tested whether including year as random factor improved the fit of the model, measured by Akaike information criterion (AIC) values. A significant main effect (P < 0.05) was further investigated by performing a Tukey’s post-hoc test using estimated marginal means with the ‘emmeans’ package in R (Lenth 2021). Test assumptions of normality of residuals (e.g. Shapiro-Wilk test) and homogeneity of variances (‘car’ package; Fox & Weisberg 2019) were checked before calculating the results. To analyse the variation in maximum numbers between sites and years, we calculated mean values, standard deviation (SD) and range (Altman & Blant 2005).
To explore whether the occurrence of juvenile individuals inside each of the three resighting areas was linked to location of their area of origin, we calculated Odds Ratios (OR) and 95% confidence intervals (CI) by use of the ‘fmsb’ package in R (Nakazawa 2021). For this, the ratio of resighted birds from one ringing area seen in one of the resighting areas to birds of other origins seen in the same area was divided by the ratio of resighted birds from the one ringing area seen in the other two resighting areas to the birds of other origin seen in the same other two areas. Individuals from one ringing location that were only seen in one of the resighting areas could not be analysed. An OR > 1 indicated a positive association between resighting and ringing area, while an OR < 1 indicated a negative and an OR = 1 indicated no association.
RESULTS
Phenology and overall distribution
The overall phenology and maximum numbers of Sandwich Terns differed between the nine study sites (Figure 2, Table 1). A general comparison showed significant differences in the timing of peak decades between study sites (F8,85.5 = 8.3, P < 0.001) with peak values occurring later in the East and Southeast. More specifically, numbers at Blåvand (SW), Agger Tange (W) and Skagen (N) peaked earlier than at Ebeltoft (E), Djursland (E) and Gedser/Rødsand (SE). Peak numbers at Fanø (SW) and Hirtshals (N) were also earlier than at Ebeltoft (E) and Gedser/Rødsand (SE; Figure 2). Including year as a random factor did not improve the model (AIC = 396.6, model without random effect: AIC = 394.9, DAIC = 1.7, P = 0.60). Thus, there was no evidence for year-to-year variation in timing of the peak decade. However, the range and standard deviation for mean peak values at any given site showed rather large variation among years (Table 1). Hence, we found large year-to-year variation in the recorded maximum numbers at many of the sites, but the timing of those peaks did not vary to any large extent. Fanø exhibited the highest mean of annual maxima and the highest ever recorded maximum of 9560 individuals.
Figure 2.
Phenology of Sandwich Terns recorded in Denmark from June until October. (A) Comparisons of timing of mean peak decades (10-days-periods) at the nine study sites. Mean values are presented including their 95% confidence interval. X-axis presents the different study sites; the y-axis presents the time when peak numbers occurred at a site. Decade 5 represents mid-July (days 11–20), decade 6 represents the last 10 days of July, decade 7 the first 10 days of August, etc. Different letters labelling the study sites mark significantly different pairwise comparisons, while identical letters mark non-significantly different pairs. Data include years from 2008 until 2020 inclusive. (B) Overall decadal phenology during post-breeding of Sandwich Terns recorded over several years at the nine study sites analysed in this project. The x-axes present the time as decades (10-days-periods); y-axes present the average numbers of Sandwich Terns counted at each study site, with standard errors. Average numbers were derived by averaging the maximum number of Sandwich Terns recorded per decade per year.

Roosting sites: characteristics and threats
The nine study sites had many geomorphological similarities. All sites have shallow water levels with sandy shorelines. The study sites differed with respect to the structures the Sandwich Terns used as roosting spots. On Fanø (SW), Blåvand (SW), Djursland (E) and Ebeltoft (E) sandbanks were preferred for roosting during low tide. During high tide, the terns rested on the beach itself. In Hirtshals (N) and Skagen (N), the tides were not strong enough to form sandbanks on which the terns could stand well, but there were temporary ponds on the beach where the terns preferred to roost, as well as on the beach itself. At Agger Tange (W), Nordmandshage (E) and Gedser (SE), the terns were mainly resting on sandy islets, away from the actual beach or path. A common feature of all these sites was that the sandbanks, sandy islands or temporary ponds on the beach provided some kind of protective roosting place for the terns. It was evident that, on Fanø (SW), Sandwich Terns standing on a sandbank close to the beach were not disturbed at all by humans passing rapidly by. However, when the tide covered the sandbanks and the terns were forced to roost directly on the beach, they were persistently disturbed by passing human traffic. Standing in the shallow water of a temporary pond on the beach appeared to have an effect similar to the one observed when the terns roosted on a sandbank. However, this was not the case when walkers approached directly towards the terns (e.g. walking towards a flock and/or stopping in front of the terns). During 19 days with observations of Sandwich Tern responses to (human) disturbances on the beach, Sandwich Terns were disturbed on average 1.3 times per hour (d/h) in 2020. Most disturbances were caused by humans passing through the flock on foot or while practising water sports (Table 3). The few instances of disturbances caused by other birds were all triggered by Great Black-backed Gulls Larus marinus. In all cases, the disturbances caused the terns to fly up briefly, although some usually landed again around the same spot. However, in most cases it was only a reduced number of the terns that returned to land again, resulting in a net displacement of birds. Examples along the Hirtshals Coast (N) showed that frequent disturbances could cause a flock to reduce noticeably in size or even disappear. For instance, during one observation day, five water sport disturbances in one hour (5 d/h) caused a flock of Sandwich Terns (n = 45) to abandon the area completely. On another day, a flock of 250 terns was reduced by 88% after experiencing 20 disturbances by human walkers in 5.5 hours (3.6 d/h).
Use of Danish waters in relation to geographical origin
In total, 1765 individual Sandwich Terns marked with colour and vertical rings were resighted in different areas in Denmark (Southwest: 1433, North: 214, East: 118; see also Table S2). Odds Ratios (OR) were used to reveal the association between resighting and ringing areas, with positive associations been symbolised by OR >1. For half of the areas of origin, the OR estimates showed a positive association between the ringing and resighting area (Table 2). Hirsholm birds (DK) were positively associated with resightings in the East and North, birds from Southeast Denmark were positively associated with resightings in the East. Birds from Baltrum (GER), Norderoog (GER) and the northwestern part of The Netherlands were associated with Southwest Denmark. All of those populations were either not or negatively associated with the other resighting areas. Sandwich Terns from West Denmark, the Delta region (NL) and Poland showed no association to any resighting area (Table 2). Because ratios were compared with each other to calculate associations, differences in numbers of observed individuals between resighting areas did not influence the results. British and Northeast-German birds were only seen in one of the resighting areas, preventing comparisons for these colonies.
Table 1.
Mean annual maximum numbers of Sandwich Terns over several years from the nine study sites. The term ‘Area’ refers to the different coastal parts of Denmark. Range and standard deviation (SD) are used as a measure of variability among years.

Sandwich Terns from different origins showed different spatiotemporal patterns (Figure 3). In Southwest Denmark, Danish and Norderoog (German northern Wadden Sea) tern numbers peaked early in the season by the end of July. However, Dutch and Baltrum (German southern Wadden Sea) birds peaked in mid-August (Figure 3A, C). In the East, birds peaked early in July, although in low numbers. Peaks in numbers from other breeding populations appeared later, generally by the end of August (Figure 3D). In the North, birds dispersing from the local colony Hirsholm (DK) already reached their maximum at the beginning of July. Most birds from Norderoog (GER) that turned up in the North were seen shortly after the end of July, while Dutch birds peaked by the end of August (Figure 3B). Sandwich Terns that originated from Poland and Great Britain are not included in the graphs, since only few individuals from these populations were seen in Denmark. However, resightings of those birds were made late in the season as well. In Southwest Denmark, peak numbers of Polish birds (n = 4) were seen in mid-September. In North and East Denmark, only one Polish individual was seen at the end of August and mid-October, respectively. The three British birds observed in Southwest Denmark were recorded at the end of July, mid-August, and mid-September. One Sandwich Tern from Northeast Germany was seen three times in East Denmark from the beginning of August until mid-September.
Table 2.
Odds Ratios (OR) calculated for juvenile Sandwich Terns that dispersed from different ringing areas to three different regions of Denmark. ORs for birds belonging to a certain ringing area were derived by comparing resightings from individuals made in the areas of either East, North or Southwest Denmark with resightings made in both other areas. For example, resightings of birds belonging to one area of origin made in East Denmark are compared with resightings of the same population made in North and Southwest Denmark. Data include years from 2011 until 2020. OR > 1 indicates a positive association between the resighting and the ringing area whereas OR < 1 indicates a negative association and OR = 1 indicates no association. A hyphen indicates that either no data were available or only for one area, so that no comparison could be made. Confidence intervals (CI) are shown on a 95% level. Significance levels: ns = not significant, *P < 0.05, **P < 0.01, ***P < 0.001.

Table 3.
Frequency of disturbances that Sandwich Terns experienced at five study sites in Denmark during post-breeding in 2020. The five study sites were Fanø, Blåvand, Hirtshals, Djursland and Ebeltoft and observations were made over 19 days. In all cases, disturbances by other birds were caused by Great Black-backed Gulls Larus marinus.

DISCUSSION
The results of this study demonstrated the importance of shallow sandy bottomed Danish inshore coastal waters as post-breeding habitat for European Sandwich Terns, with marked differences in temporal and dispersal patterns between east and west coasts. Dispersing marked juveniles from adjacent European countries showed that the location of their fledging colonies influenced their use of Danish post-breeding roosting sites and phenology and the critical international importance of Danish waters during post-breeding dispersal for the species. Levels of human disturbance to resting Sandwich Terns confirmed the need for protected roosting areas on beaches with intense human activity.
Spatiotemporal distributions
Our data show that Sandwich Terns use Danish waters during post-breeding mainly between the end of July and the beginning of September. Maximum numbers occur in August, culminating in the beginning of August along the western part of Denmark (here including the study sites Southwest, West and North Denmark) and along the eastern part of Denmark (the study sites East and Southeast Denmark) during mid-August and September. The overall phenology and timing of peak numbers in Denmark are rather similar to the post-breeding phenology in other areas of northern Europe. For example, in England peak numbers of roosting Sandwich Terns are recorded in mid-August and in Ireland between mid and end of August (Ward 2000, Burke et al. 2020). In Poland, Sandwich Terns show a bimodal peak: in late July (comprising local breeding birds) and again mid-August until September (Meissner 1999). On the Iberian Peninsula, maximum numbers of migrating terns pass by in September (Fernández-Cordeiro & Costas 1991), slightly later than the maxima recorded at northerly post-breeding areas.
Figure 3.
Temporal variation in the occurrence of juvenile Sandwich Terns originating from different breeding areas located in Denmark as well as in neighbouring countries. Note that both plot A and C refer to the resighting area in Southwest Denmark from different breeding areas. The x-axis present time in 10-day-intervals (decades) from June until October; y-axis presents numbers of individuals. Individuals can contribute multiple observations in different decades. Different colours show different areas of origin. Data include years from 2011 until 2020 inclusive.

The results show that the nine roosting sites that were analysed in this study differed in local abundance, with Fanø (SW) having by far the highest numbers of roosting terns. Mean maximum values from Fanø (SW) exceed the 1%-threshold criteria which qualifies as being internationally important as a roosting site for European Sandwich Terns (Population 1%-level = 1700; Wetlands International 2021). However, range values for mean maxima for Blåvand (SW), Agger Tange (W), Skagen (N) and Nordmandshage (E) showed that those sites also qualify as being sites of international importance. Furthermore, the results confirm, that numbers can vary markedly between years and all study sites showed large variations in annual maximum number, findings similar to those from Ireland (Burke et al. 2020).
It appears that certain sites along the west and northeast coast of Jutland are unique in terms of the numbers of Sandwich Terns that – in most years – use the area during the post-breeding period. We have not been able to find information on more than a handful of sites around the North Sea and the Baltic Sea that regularly have roosting flocks of more than 300 Sandwich Terns during July–September.
Drivers of spatiotemporal patterns
Various factors are likely to influence the spatiotemporal patterns that characterize the post-breeding distribution of dispersing juvenile Sandwich Terns with their parents. We predict an aggregative response of Sandwich Terns to high local abundance of suitably sized fish prey, as the terns will need energy resources to accomplish their migratory journey to the wintering areas along the coast of Africa. The availability of disturbance-free roosting sites, combined with access to alternative roosting sites in case of disturbance, could influence distribution patterns as well. Proximity to safe roosting spots and rich feeding grounds are likely to be important factors since this would lead to short foraging trips, fewer disturbance flights and thereby maximize the net energy uptake. It is especially important for juveniles to have safe roosts while waiting for their parents to feed them. Moreover, the parents need to know where their offspring are located in order to efficiently deliver food. Thus, connectivity of safe roosting spots is likely to be important as well. Furthermore, the proximity between breeding colonies and post-breeding roosting sites might influence the spatiotemporal patterns. The first roosting site to which a tern arrives depends on its location with respect to its breeding colony. To save energy, it might be advantageous to stay at the first best foraging and roosting site it encounters. More factors could affect the spatiotemporal distribution patterns. These include wind force and wind direction that might influence fish distribution, the detectability of fish as well as the size of roosting spots, e.g. the width of the beaches on which the terns can roost (including access to alternative nearby sites in case of disturbance). In the following, the three first mentioned factors (prey availability, disturbance-free roosting sites and the proximity between breeding and post-breeding site) will be discussed in more detail.
One possible explanation for different occurrences of maximum numbers of Sandwich Terns along Danish waters could be the timing and distribution of their key prey species, thought to be sandeel, Sprat and herring. All three fish species are highly abundant and widely distributed in Denmark, albeit with large local variation, and utilise shallow coastal waters as nursery areas for their juvenile and one-year-old fish (Hoffmann & Carl 2019, Munk et al. 2019, Munk & Carl 2019, Møller et al. 2019), on which terns mainly forage (Cabot & Nisbet 2013). Sandeels are particularly abundant in Danish Waters and different Ammodytes species are present in the North Sea, Kattegat and Baltic Sea. However, the highest abundance of sandeels can be found in the North Sea (Møller et al. 2019, Munk et al. 2019). Reasons for this include the tidal influence in that area, especially in the Wadden Sea, which provides abundant oxygen and sand supply (Wright et al. 2000, Munk et al. 2019), an important factor for sandeels as they exhibit a distinctive burying behaviour, which they employ to sleep during the night but also to overwinter for several months in the sandy seabed (van Deurs et al. 2011, Munk et al. 2019, Møller et al. 2019). Horns Rev, a reef off the Danish Wadden Sea close to Blåvand (SW) and Fanø (SW), is a particularly important habitat for sandeels, supporting several species in high densities (Munk et al. 2019, M. van Deurs pers. comm.). Fisker Bank, a sand bank off the Danish west coast (close to Agger Tange (W)) is another place with suitable sandeel habitat, supporting high densities (Munk et al. 2019). Furthermore, the results of this study show, that high numbers of roosting Sandwich Terns are often found along the west coast of Denmark, close to the suitable sandeel habitats. However, sandeels, that are at the right size for Sandwich Terns to feed on, overwinter buried in sand for eight months from approximately August/September until March/April (Winslade 1974, Munk et al. 2019, Møller et al. 2019) and thus start to disappear during the main phase of post-breeding dispersal of young birds and their parents. Nevertheless, before they start to overwinter, high densities of sandeels aggregate in the wintering habitats in the Danish North Sea region, which is therefore likely to become more attractive to feeding Sandwich Terns at that time than the Danish Kattegat region. This could explain why the study sites along the Danish west coast (i.e. the North Sea) peaked in the beginning of August as the Sandwich Terns seem to adjust their distribution to the timing and phenology of their prey. After sandeels start overwintering, Sandwich Terns might redistribute themselves more evenly among the study sites to forage on Sprat and herring, which are still abundant in the Danish North Sea and Kattegat in August and September (ICES 2005, Munk & Carl 2019). Especially in Skagerrak and Kattegat, larger numbers of Sprat aggregate, which could explain why the study sites in the east (i.e. the Danish Kattegat and Baltic Sea) peaked mainly by the mid/end of August and beginning of September after sandeels became less abundant. However, peak numbers there were still lower compared to numbers in Southwest Denmark, making the latter region most significant for Sandwich Terns.
Disturbance rates can also be expected to influence the selection of post-breeding roosting sites of Sandwich Terns. Additionally, an explanation for why long stretches of the Danish west coast were not used exclusively until sandeels started to overwinter might be the rarity of disturbance-free roosting spots in close proximity to feeding grounds. Especially during the summer months, the Danish Wadden Sea as well as the west coast is characterised by high levels of tourist activity. A previous study by van der Kolk et al. (2021) has shown inverse correlations between roosting numbers of Eurasian Oystercatchers Haematopus ostralegus and human disturbances in the Dutch Wadden Sea. Similar results found in our study show that Sandwich Terns at some sites experienced more than one disturbance per hour on average and that frequent disturbances could cause a flock to abandon a roosting site completely. It is important for terns to have alternative roosting places between which they can switch in case of being disturbed. For example, cars are permitted to drive on the beach on Fanø (SW) and along the coast of Hirtshals (N), which attracts many tourists and therefore increases disturbance rates. On Fanø (SW), cars are only allowed in the middle and southern part, while the north end of the island remains car-free. Field observations showed that Sandwich Terns in southern Fanø (SW) which experience more disturbances due to a higher number of tourists, roost in lower numbers (or abandon the area) and have a higher flight initiation distance than on the north end of the island (K. Fischer unpubl. data). Along the coast of Hirtshals (N), Sandwich Terns were able to switch between alternative roosting spots. Ringed birds that were previously disturbed could often be resighted in a temporary pond, where cars could not drive close by. However, these alternative roosting sites were not completely disturbance-free and the terns often needed to shift between roosting spots.
A third explanation for the differences in occurrence of maximum numbers could be the connectivity (i.e. the outline of coasts) and the distance between the breeding colonies and the different parts of the Danish waters that the terns exploited post-breeding. For example, Sandwich Terns may disperse along the coastlines until they encounter the first suitable site that provides resources in terms of food and safe roosting places. The results of this study support the hypothesis that terns from different breeding populations were associated with different post-breeding areas in Denmark. They show that the North Sea populations of Sandwich Terns were indeed associated with roosting sites along the Danish west coast that were closest to their breeding/hatching colony (particular on Fanø (SW)) and resightings of birds ringed in breeding colonies in the Danish Kattegat showed an association with areas in North and East Denmark. Furthermore, numbers of breeding pairs in the North Sea region, e.g. The Netherlands, are much higher than numbers of breeding pairs in the Baltic region, e.g. Poland (see Buijsman 2020), which could explain differences in roosting numbers along the western and eastern part of Denmark. Although Polish birds were seen in all of the three resighting areas (albeit in low numbers), they showed no association to any area. However, due to lack of breeding success at the Polish colony since 2016, no chicks have been ringed in Poland since then (S. Bzoma pers. comm.). Furthermore, no juvenile birds with colour- or vertical rings of Swedish origin have been observed in Danish waters, and only a single colour-ringed juvenile from Northeast Germany was observed in Denmark, making it impossible to describe the patterns of post-breeding dispersal of juveniles with their parents originating from the Baltic region. The patterns of phenology based on resightings of marked individuals from different breeding colonies suggests that in general, birds from more distant breeding colonies turn up later than birds originating from breeding colonies located closer to the post-breeding areas in Denmark. For example, most resightings of Dutch birds occurred later in Southwest Denmark than birds dispersing from closer colonies. Dutch birds also peaked late in North and East Denmark, albeit in much lower numbers. These birds may have previously exploited rich feeding areas that are closer to their colony, as most Dutch birds leave their colonies by the end of June and beginning of July (R. Fijn pers. comm.). Polish birds were seen late in the season, which suggests a similar pattern.
Based on the results of this study, it seems probable that the distribution and phenology of fish, in combination with availability of safe roosting spots as well as the distance to the colony of origin are important drivers for the spatiotemporal patterns in the post-breeding distribution of Sandwich Terns in Denmark. However, more data from Baltic colonies like Poland, Sweden and Northeast Germany would be useful to improve our understanding of the connection between the location of breeding colonies and the timing of occurrence of individuals at different potential post-breeding areas. Due to the low number of chicks ringed with vertical metal rings or colour-rings in the Baltic Sea, it also remains unclear to what extent late arrival of individuals from distant Baltic breeding colonies could help to explain the later peak in numbers along the easterly Danish coasts.
Variations in locally roosting numbers between years could also be explained by variation in local fish abundance. Population sizes of fish are highly variable between years, mainly due to variations in mortality and recruitment, which affects annual transition of age groups to the next life stage (Daan et al. 1990, Camp et al. 2020, ICES 2021a,b,c,d). Differences in breeding success of Sandwich Tern colonies between years could be another explanation. Bearing in mind that it takes a long time for the juveniles to hone their feeding skills and that the parents usually do not stop feeding their offspring before they reach the wintering area (Dunn 1971, Cabot & Nisbet 2013), failed breeders could be expected to spend less time in potential post-breeding areas and instead migrate to Africa earlier than families.
Limitations of sampled data
Volunteers collected most of the data analysed in this study, and this had advantages and disadvantages. For example, without all the volunteer data entries in DOFbasen over several years, it would not have been possible to gather such an extensive dataset or to generate a nationwide overview of the distribution and abundance of post-breeding juvenile Sandwich Terns and their parents in Denmark. However, the effort differed greatly between locations and sites. For example, Fanø (SW), Blåvand (SW) and Skagen (N) had the highest and most standardised effort, and data from many years could be included in this study, whereas the other sites were counted in less standardized ways. For all nine study sites, there were years when no Sandwich Terns were counted in some of the decades. However, for almost all gaps it was unknown whether Sandwich Terns were absent and therefore not reported or whether the site was not covered and thus terns that were present were not counted. It might also be possible that some important sites were not counted because they were not known/accessible to observers. However, the indication that counts at Gedser (SE) were able to reflect roosting numbers on Rødsand makes it likely that most of the important sites, even those that were not accessible to observers, were discovered. Additionally, the exact timing of an observation may have influenced the number of counted birds. For example, less birds might have been counted when an observer arrived shortly after a disturbance or when many of the terns were out foraging at sea and were therefore not seen. The same observer-effort applies to resightings of individually marked birds with different ring types that are likely to have different detection and reading probabilities. It is unknown how thoroughly observers scanned the roosting flock to find ringed individuals. Furthermore, it is likely that ringing effort in colonies as well as resighting efforts at different areas in Denmark varied between years. Also, the results in relation to geographical origin can only reflect phenology patterns of families, but not of adults without offspring. Thus, all counts and numbers of resightings of Sandwich Terns presented in this study can only be considered as minimum numbers. Additionally, data on disturbance rates were only collected in 2020 and more surveys from different years are needed to provide more robust data.
Conclusions and future prospects
This study highlights the international importance of Danish waters as a post-breeding area for Sandwich Terns breeding in the international Wadden Sea as well as in the Baltic Sea. Our study showed that a number of Sandwich Tern families from colonies located in the Dutch and especially in the German part of the Wadden Sea moved north to Danish waters post-breeding, apparently because these waters offered good opportunities for fattening up before the autumn migration to west and southern Africa. Danish waters are also used during post-breeding by families of Baltic origin, and these families can select and stopover in areas located en route towards the wintering areas. Many of the attractive roosting areas in Denmark were characterized by having multiple roosting spots (birds could move short distances when disturbed) and were in proximity to feeding areas offering a high abundance of fish (see Burke et al. 2020). There was some evidence that the terns within and between seasons adapt their choice of site to the spatio-temporal variation in the availability of their key prey species. To better understand post-breeding distribution patterns of Sandwich Terns across Europe, more studies of roosting numbers and movement patterns are needed.
Future studies should concentrate on experimentation with conservation measures, with particular emphasis upon the precise location, size and structure of disturbance free roosting sites on beaches (see van der Kolk et al. 2021), in order to protect this species outside the breeding season. Knowledge about prey choice and monitoring of food abundance and availability throughout the season around important roosting sites could help to understand dynamics and differences in annual numbers better. Additionally, the use of GPS transmitters concentrating on the period of post-breeding of European Sandwich Terns from various colonies could provide detailed information on how dynamic individuals are in their movements among potential post-breeding sites and possibly reveal differences between families and single individuals in the use of these sites.
ACKNOWLEDGEMENTS
Special thanks go to Thomas Alerstam, who enriched the analysis and the manuscript with helpful comments and suggestions throughout the process. We would like to thank all voluntary observers that contributed to this project by reporting count numbers and readings of individually marked Sandwich Terns. Likewise, a special thanks goes to Skagen Bird Observatory and Blåvand Bird Observatory. We thank Kjeld T. Pedersen and the Danish Ringing Centre for providing various information of resighted Sandwich Terns. From Germany, Ulrich Knief (University of Munich), Matthias Haupt, Olaf Geiter (Vogelwarte Helgoland) and Christof Herrmann (Vogelwarte Hiddensee) contributed with delivering life-histories of Sandwich Terns seen in Denmark. From The Netherlands Ruben Fijn (Bureau Waardenburg, Netherlands), Wouter Courtens (Research Institute Nature and Forest, Belgium), Deltamilieu Projecten (Netherlands), Mardik Leopold (Wageningen Marine Research), Allix Brenninkmeijer and Date Lutterop contributed with similar data and helpful discussions. Bridget Griffin (British Trust for Ornithology) and Szymon Bzoma (University of Gdańsk) provided information of birds ringed in Britian and Poland, respectively. Thorsten J. Skovbjerg Balsby and Jessica Abbott helped to improve the statistical analysis. We want to thank Anna Rindorf and Mikael van Deurs for providing information about the complex life histories of fish.
REFERENCES
Appendices
SAMENVATTING
Grote Sterns Thalasseus sandvicensis verspreiden zich na het broedseizoen, net als andere soorten sterns, over enige afstand rond de broedplaats (dispersie) voordat zij naar hun overwinteringsgebied trekken. Wij hebben aan de hand van 2422 waarnemingen uit eind juni tot oktober 2011–2020 van individueel met kleurringen gemerkte vogels onderzocht wat de betekenis van de Deense wateren in dit dispersieproces is. Naast eigen waarnemingen hebben wij ook gebruik gemaakt van gegevens uit de online database van de Deense Ornithologische Vereniging (DOFbasen). Daarnaast hebben wij de habitatstructuur en de mate van menselijke verstoring van de belangrijkste rustplaatsen van de sterns in het veld vastgelegd. Uit het onderzoek is gebleken dat de vogels in het westen van Denemarken eerder in de tijd de grootste aantallen bereiken dan in het oosten van het land. De belangrijkste rustgebieden voor sterns worden gekenmerkt door zandstranden met ondiepe kustwateren, een weerspiegeling van de habitat van hun belangrijkste prooidieren. De resultaten van het onderzoek suggereren dat de ligging van de rustgebieden en de bezetting van die gebieden worden bepaald door de nabijheid van de broedkolonies. Het grote aantal sterns uit andere landen dat werd waargenomen, geeft aan dat de Deense wateren tijdens de dispersieperiode van grote internationale betekenis zijn voor de soort. Menselijke verstoring van rustgebieden trad geregeld op (tot gemiddeld meer dan één verstoring per uur) en onderstreept de noodzaak van bescherming van stranden met veel menselijke activiteit.
SUPPLEMENTARY MATERIAL
Table S1.
Showing the analysed years per site.

Table S2.
Numbers of Sandwich Terns ringed as pulli and resighted as fledged juveniles in different areas in Denmark between 2011 and 2020 by ring type. Note, that the number of ringed individuals does not provide any information on how many chicks have fledged and survived during their first calendar year.