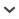A new species belonging to the recently described genus Malagodon Meinema and Huber, 2023, is herein described from specimens collected in a single, small, isolated Pandanas swamp in southeastern coastal Madagascar, located within the Réserve Spéciale de Manombo, south of the town of Farafangana, its only known locality. The new species was last collected in the late 1990s, and despite repeated attempts over the past three decades, no additional specimens have been collected at the type locality or from any other suitable habitats within the region, and the species is presumed to be extinct. The new species is distinguished from its only congener, Malagodon madagascariensis, formerly known from forested swamps in northeastern coastal Madagascar, and also now considered to be extinct, by the following apomorphic features: a lower anal-fin ray count (15–17 vs. 18–19), a longer caudal peduncle (26.8%–39.8% vs. 21.9%–26.7% SL), and the presence of a platelike (vs. thin and spinelike) neural spine on the fifth vertebral centrum in both sexes. Additionally, the new species exhibits neural spines on both the sixth and seventh vertebral centra that are also somewhat expanded and platelike dorsally compared with those in M. madagascariensis, which are narrow and spinelike. We also provide a rediagnosis of Malagodon based on the examination of additional material unavailable in the original description, which was based on only three specimens (two males of M. madagascariensis and one female of the new species).
INTRODUCTION
Members of Malagodon Meinema and Huber, 2023, are small oviparous cyprinodontiform fishes (<30 mm SL, usually considerably smaller) that formerly inhabited forested coastal swamps in eastern Madagascar. The only species in this currently monotypic genus, M. madagascariensis, was endemic to forested swamps in northeastern coastal Madagascar near Mahambo (17° 29′ S, 49° 28′ E), located to the north of the major port city Toamasina (Tamatave), and is now considered extinct (fig. 1). A second species, known only from a single, small, isolated Pandanas (screw pine or screw palm) swamp in southeastern coastal Madagascar, located within the Réserve Spéciale de Manombo (23° 3′ S, 47° 42′ E), south of the town of Farafangana, is herein formally described (fig. 1). The new species was first discovered in 1994, last collected in 1997, and is also currently presumed to be extinct (Ravelomanana et al., 2018; IUCN, 2023; Sparks and Smith, 2022). The two species of Malagodon are known only from their type localities, which are separated by nearly 650 km (400 miles), and despite targeted efforts, no individuals have been collected from presumably suitable habitat in the intervening region.
Prior to the work of Meninema and Huber (2023), Malagodon madagascariensis was ascribed to Pantanodon, a monotypic genus described by Myers (1955) to encompass the morphologically distinct species P. podoxys. In the following decades, two additional species were ascribed to Pantanodon Myers, 1955, such that until the mid 1990s the genus contained three species that were considered valid: P. stuhlmanni (Ahl, 1924), P. podoxys Myers, 1955, and P. madagascariensis (Arnoult, 1963). Pantanodon podoxys was subsequently determined to be a junior synonym of Haplochilichthys stuhlmanni (Seegers, 1996). Therefore, up until the study of Meinema and Huber (2023), Pantanodon comprised only two valid species, P. stuhlmanni, ranging from Kenya to Tanzania, and P. madagascariensis, formerly endemic to coastal swamps in northeastern Madagascar.
In his brief description of P. podoxys, Myers (1955) created the monotypic subfamily Pantanodontinae within Cyprinodontidae exclusively for this morphologically distinct species. This taxonomic arrangement was supported by Whitehead (1962), who likewise treated Pantanodontinae as a subfamily of Cyprinodontidae. Rosen (1965) subsequently redescribed Oryzias madagascariensis (Arnoult, 1963) and assigned this Malagasy taxon to Pantanodon based on several anatomical features he considered unique to P. podoxys and P. madagascariensis at the time (see Rosen, 1965: 5–6). Based on these features, Rosen (1965) recommended classifying the genus in its own family, Pantanodontidae, because its immediate relationships remained unclear. Clausen (1967) further suggested that Pantanodon may be closely related to modern aplocheilichthyine poeciliids. In the first comprehensive phylogenetic analysis of Cyprinodontiformes, Parenti (1981) presented a considerably revised phylogeny and classification of the order based on morphological characters, corroborating Clausen's hypothesis, and placed Pantanodon within Poeciliidae, subfamily Aplocheilichthyinae, which was later supported by Ghedotti (2000). In contrast to explicit morphological studies, recent multigene molecular studies have recovered Pantanodon (represented only by the African species, P. stuhlmanni) in its own family that is recovered as sister to Cyprinodontoidei and only distantly related to Poeciliidae and Aplocheilichthyinae (Pohl et al., 2015; Bragança et al., 2018). Preliminary genomic analyses using ultra-conserved elements that included P. stuhlmanni and the novel undescribed Malagasy species corroborate the monophyly of a transcontinental Pantanodon, the recognition of Pantanodontidae as a distinct family, and the placement of Pantanodontidae as sister to all other cyprinodontoids (Sparks and Smith, 2022; Sparks, Davis, and Smith, unpublished data).
FIGURE 1.
Map of Madagascar showing type localities of Malagodon honahona, new species, small Pandanas swamp, southeastern coastal Madagascar, located within Réserve Spéciale de Manombo, south of Farafangana, and M. madagascariensis, forested swamps, northeastern coastal Madagascar near Mahambo, north of major port city Toamasina (Tamatave).

Although the immediate relationships and composition of Pantanodon and related taxa have been problematic and remain somewhat unresolved (Sparks and Smith, 2022), Meinema and Huber (2023) provide a detailed and much needed taxonomic review and update of both recent and fossil forms, and recognize an expanded Pantanodontidae comprising four genera, two of which they described as new: Pantanodon (two extant species from coastal Tanzania); Aliteranodon Meinema and Huber 2023 (five extant species, plus an undescribed form, all from coastal Kenya and Tanzania); Malagodon Meinema and Huber, 2023 (one extinct species and one undescribed taxon, both formerly endemic to eastern coastal Madagascar); and Paralebias, comprising all extinct fossil European species. We note that Paralebias is not currently diagnosed by apomorphic features, and that it may not comprise a monophyletic assemblage (Bragança et al., 2018).
Meinema and Huber (2023) placed their newly described monotypic genus, Malagodon, which included only P. madagascariensis, as the sister taxon to Pantanodon. Species of Malagodon historically exhibited a very restricted distribution within Madagascar, endemic to forested coastal swamps in eastern Madagascar, whereas the range of their reconfigured Pantanodon is restricted to Tanzania. Specifically, P. podoxys, removed from synonymy with P. stuhlmanni, is now confined to southern coastal swamps and lagoons in Tanzania, and a newly described species, P. propinquus Meinema and Huber, 2023, is restricted to northern coastal swamps and lagoons in the Tanga region of Tanzania. Meinema and Huber (2023) also revalidate P. stuhlmanni (Ahl, 1924), propose its type locality as Pangani (its only known locality) in northeastern Tanzania, and place this taxon in their newly described genus Aliteranodon, along with four other newly described species and one undescribed taxon, all from coastal Tanzania and Kenya (see Meinema and Huber, 2023: fig. 2). Thus, in this significant taxonomic reorganization, two of the three Recent genera comprising Pantanodontidae, Aliteranodon and Pantanodon, are restricted to East Africa, with only Malagodon endemic to Madagascar.
Although Meinema and Huber (2023) provide a rather lengthy diagnosis for Malagodon, we provide a rediagnosis below that is restricted to the primary apomorphic features that separate Malagodon from all other Recent members of mainland African Pantanodontidae. Malagodon were small, colorful fishes that inhabited extremely shallow water above a thick turbid layer of sediment in forested brackish swamps. Due to habitat destruction and deforestation, it is assumed that all members of Malagodon are now extinct (Sparks and Smith, 2022). Whereas we agree with the creation of Malagodon to encompass the Malagasy members of Pantanodontidae, the description of the genus by Meinema and Huber (2023) was based on only three specimens of Malagodon, two males of M. madagascariensis and one female of the new species. Thus, a redescription of the genus based on additional material is warranted. Herein we describe a new species of Malagodon from southeastern coastal Madagascar and rediagnose the genus based on the examination of numerous additional specimens of both species.
MATERIALS AND METHODS
Specimens used in this study were fixed in 10% buffered formalin and stored in 75% ethanol. Cleared and double-stained skeletal preparations (C&S) were produced using a protocol based on Taylor and Van Dyke (1985). Counts and morphometric measurements follow Barel et al. (1977), Hubbs et al. (2004), and Sparks (2008), unless noted otherwise. All morphometric measurements were recorded to the nearest 0.1 mm. Measurements were verified at least twice using analog dial calipers. Vertebral and fin ray counts were obtained from radiographs and C&S skeletal preparations. Vertebral counts exclude the terminal, hypural-bearing vertebra (= terminal half centrum). The first caudal vertebra is defined as the first vertebra bearing a definitive hemal spine (Hubbs et al., 2004), where the hemal spine comprises greater than half of the ventral vertebral anatomy. The terminal dorsal- and anal-fin rays are counted as a single element, even if branched and split to the fin base, as this element is associated with a single supporting pterygiophore. Pored scales in the lateral line are counted in series from the dorsal margin of the gill opening to the caudal flexure. Pored scales posterior to the hypural flexion are excluded from the count, regardless of size. Osteological features of the new species and related taxa were examined using alcohol-preserved specimens, C&S skeletal preparations, or obtained from radiographs. Gill-raker counts (i.e., lower limb of the first gill arch) exclude the raker in the angle of the arch marking the transition from ceratobranchial to epibranchial. To determine significant differences between the two Malagodon species for various morphometric measurements, a Shapiro-Wilk Normality Test (Shapiro and Wilk, 1965) was performed for each measurement with the “stats” package in R (version 4.3.0; R Core Team, 2023). An analysis of variance (ANOVA) and Tukey HSD (Tukey, 1949) or Kruskal-Wallis (Kruskal and Wallis, 1952) and Pairwise Wilcoxon Rank Sum (Wilcoxon, 1945) tests were then performed for parametric or nonparametric data, respectively with a 95% confidence interval. Note that all specimens of the new species could not be included in the data table for all counts/measurements due to poor preservation/damage in a few individuals. Institutional abbreviations are as listed in Leviton et al. (1985) and Sabaj (2020; 2023).
RESULTS
Systematic Account
Malagodon
Figures 2–7
Diagnosis: The primary apomorphic features that separate Malagodon from all other Recent members of mainland African Pantanodontidae are: the presence of oral (external) jaw teeth in both sexes; a pleural rib count of 12–14; the pelvic fins in males are highly modified, with up to the first three pelvic rays becoming notably thickened and spinelike in larger individuals, including the presence of paired claws and hooks on pelvic rays 1–4 (i.e., number of spinelike processes increases, and hooks continue to develop on additional (posterior) pelvic rays as the individual matures; see progression with ontogeny in fig. 6); fully fused hypural plates in the caudal skeleton; and both the dorsal and anal fin of males are notably elongated and produced, with long trailing margins in larger individuals.
Malagodon honahona, new species
Figures 2–6; table 1
Pantanodon n. sp. “manombo”: Sparks and Stiassny, 2003: table 9.1.
Pantanodon n. sp. “manombo”: Sparks and Stiassny, 2008: table 1.
Pantanodon n. sp. “manombo”: Sparks and Stiassny, 2022: tables 10-1 and 10-9.
Pantanodon n. sp. “manombo”: Sparks and Smith, 2022.
Malagodon spec. ‘Manombo Reserve’ (yet undescribed): Meinema and Huber, 2023.
Holotype: UMMZ 240245; male, 23.9 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 94-4; P. Reinthal, J. Sparks, and K. Riseng, 16 Jun. 1994.
Allotype: UMMZ 254162; female, 18.9 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 97-2; P. Reinthal et al., Oct. 1997.
Paratypes: AMNH 278989 (formerly UMMZ 240245); 2 ex. C&S (1 male and 1 female), 18.6–20.2 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 94-4; P. Reinthal, J. Sparks, and K. Riseng, 16 Jun. 1994. AMNH 278990 (formerly UMMZ 254161); 2 ex., 20.4–20.7 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 97-2; P. Reinthal et al., Oct. 1997. UMMZ 254159 (formerly UMMZ 240245); 2 ex., 19.4–20.2 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 94-4; P. Reinthal, J. Sparks, and K. Riseng, 16 Jun. 1994. UMMZ 254160; 5 ex., 16.2–23.2 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 94-3; P. Reinthal, J. Sparks, and K. Riseng, 15 Jun. 1994. UMMZ 254161, 13 ex., 15.3–21.8 mm SL; Madagascar: Fianarantsoa Province: Réserve Spéciale de Manombo; small Pandanas swamp at edge of intact forest; 23° 3′ S, 47° 42′ E; PNR 97-2; P. Reinthal et al., Oct. 1997.
Diagnosis: The new species is readily distinguished from Malagodon madagascariensis, its only congener, by the following apomorphic features: lower anal-fin ray count (15–17 vs. 18–19), longer caudal peduncle (26.8–39.8 vs. 21.9–26.7% SL), and notably expanded and platelike neural spine on arch of fifth vertebral centrum in both sexes (vs. thin and spinelike in M. madagascariensis). In addition, neural spines on both sixth and seventh vertebral centra in new species also somewhat expanded and platelike dorsally versus narrow and spinelike in P. madagascariensis.
Description: Morphometric and meristic data presented in table 1. Pigmentation pattern in life and preservation, as well as external and internal morphological features, shown in figures 2–6. Body similar in overall shape to M. madagascariensis, and relatively elongate and dorsoventrally flattened anteriorly. Body width of males becoming gradually more laterally compressed posteriorly, whereas body width of females becoming significantly laterally compressed posterior to pelvic fins. Head extremely dorsoventrally flattened. Snout pointed and oriented dorsally. Orbit very large. Eyes dorsolaterally oriented. Interorbital width approximately half of orbit diameter. Mouth wide, superiorly oriented, and jaws short. Posterior margin of maxilla extending posteroventrally to vertical approximately at midpoint of orbit. Upper lip large and thickened medially. Lower lip thin. Lower jaw greatly protrudes anterior to upper jaw.
Three rows of sharp lingually curved villiform teeth restricted to the premaxilla, becoming reduced to a single row posterolaterally. Tooth size decreasing laterally from premaxillary symphysis. Lower jaw with two distinct rows of lingually curved villiform teeth at symphysis of dentary, slightly decreasing in size posterolaterally. Paired 5th ceratobranchial bones (IF = inferior pharyngobranchials of Rosen, 1965: fig. 3) with 10 rows of sharp, slightly posteriorly recurved villiform teeth. Tooth number decreasing anteriorly with 12–13 teeth in the most posterior row decreasing to one tooth in the most anterior row. Pharyngobranchials 3,4 (PB3,4 of Rosen 1965: fig. 3) with eight rows of sharp, slightly posteriorly recurved villiform teeth. Tooth number greatest in medial rows, with 7–8 teeth, decreasing anteriorly and posteriorly to 3–4 teeth per row. Second pharyngobranchial elements (PB2? of Rosen, 1965: fig. 3) weakly ossified, large and elongate, tapering to a point rostrally, and with numerous associated toothplates (many weakly associated and floating in adjacent tissue as opposed to directly fused to PB2? bones).
Suborbital series comprised of the lacrimal and the dermosphenotic. Posterior margin of preopercle smooth, lacking serrations. Opercle lacking spines. Operculum extending posterior to pectoral-fin base. Gill membrane joined to isthmus, but sides meet without a connecting membrane. Lower limb of first gill arch with two rows (one inner and one outer) of 32–35 elongate, triangular rakers. Lower limb of gill arches two through four with inner and outer row rakers of a similar morphology and count (32–35 in each row) as first arch. Five branchiostegals present.
All scales cycloid, relatively uniform in size, and covering entire body. Smaller scales on caudal fin extending approximately 1/6 length of fin. Scales on nape laterally oblong. Cheek and ventrum fully scaled. Lacrimal asquamate. Snout and occiput fully scaled. Fourteen scales in diagonal from dorsal margin of opercle to origin of dorsal fin. Twenty-two predorsal scales along dorsal midline. Twenty-eight to 29 lateral line scales present.
Fin ray counts as follows: dorsal 7–8; anal 15–17; pectoral 8–9; pelvic 6; caudal 25–27. Anal fin frequently with one or two feeble leading spinelike unbranched rays (rarely three), especially in larger individuals. Pelvic fin with hooked spinelike processes (thickened rays) present in males, with both number of spinelike processes and associated hooks variable and generally increasing over ontogeny (i.e., sexually mature males generally have both more thickened spinelike rays and associated hooks; fig. 6A, C, E): (A) no spinelike process, first ray with single hook; (C) one spinelike process present, exhibiting single hook, and following ray with paired hooks; (E) two spinelike processes present, with first exhibiting single hook, second with paired hooks, and third ray with paired hooks. All females lack hooks on pelvic-fin rays regardless of size or sexual maturity. Vertebral count 17 precaudal + 13–15 caudal = 30–32 total vertebrae. Anteriormost precaudal vertebra lacking pleural ribs. Pectoral fin extending posteriorly to near vertical of longest pelvic-fin ray when both adducted. Anterior origin of dorsal fin at approximate midpoint of anal fin. Anal fin originating at vertical greater than three-quarters of predorsal distance and terminating slightly posterior to last dorsal-fin ray. Caudal fin evenly rounded to lanceolate in larger males (fig. 4).
Neural spines (= processes) notably expanded and platelike on first five vertebral centra in both sexes, not thin and spinelike (fig. 5A, B). Additionally, neural spines on both sixth and seventh vertebral centra somewhat expanded and platelike dorsally.
PIGMENTATION IN LIFE: Coloration of recently collected male shown in figure 4. Base body color orangish-brown. Head with a purplish-blue blotch that continues posteriorly as thin stripe along midline to caudal peduncle. Reddish-orange spots present on snout and bases of both pectoral and caudal fins. Unpaired fins, especially anal and caudal, reddish orange with darker margins.
Pigmentation in Preservative: Coloration in alcohol shown in figures 2 and 3. Overall coloration light grayish brown with very small dark spots covering body and visible only under microscope.
Distribution and Habitat: The new species is known only from a single, small, isolated Pandanas swamp in southeastern coastal Madagascar, located within the Réserve Spéciale de Manombo (23° 3′ S, 47° 42′ E), south of the coastal town of Farafangana (fig. 1). Although the species was first collected in 1994, it has not been collected since the late 1990s despite repeated attempts and is presumed to be extinct. The habitat was a small, turbid swamp, with only a thin layer of clear water over a much more extensive highly sedimented layer. All specimens of M. honahona were collected from this thin top layer of clear water.
Members of Malagodon are omnivorous. In aquaria, M. honahona fed on algae, zooplankton, and chironomid larvae. These fishes are oviparous, and fertilization is external. This species was kept in aquaria for several years (at UMMZ). However, efforts to induce reproduction failed. Little else is known regarding reproductive behavior of either species of Malagodon.
Conservation Status: Based on the most recent Red List assessment (IUCN, 2023), Malagodon madagascariensis is considered to be extinct (Ravelomanana et al., 2018; Sparks and Smith, 2022). Although the newly described species from southeastern coastal Madagascar was not assessed by IUCN in 2016 (Sparks, 2016), which limits assessments to formally described species, extensive collecting efforts over the past three decades by our research team and others have failed to locate additional individuals, therefore, it can be presumed that M. honahona is also extinct. Both species of Malagodon exhibited extremely restricted ranges in eastern coastal Madagascar. Malagodon madagascariensis was known only from forested areas in the region of Mahambo (17° 29′ S, 49° 28′ E), about 80 km north of the port city of Toamasina (Tamatave), and no additional specimens have been collected from the region since collection of the type series in late 1962 by Arnoult (figs. 1, 7). Malagodon honahona was first collected in 1994 by our research team from a single, small lowland Pandanus swamp within the Réserve Spéciale de Manombo (23° 3′ S, 47° 42′ E), located to the south of Farafangana, its only known locality (figs. 1, 2, 4). Unfortunately, introduced Gambusia were prevalent at this locality and likely competing with M. honahona, and specimens of the new species of Malagodon have not been collected at the type locality since 1997. The region upstream of their only known habitat lies outside the Réserve Spéciale de Manombo protected area and is afforded no protection. As a result, the watershed has experienced rapid deforestation in recent decades such that the fragile type locality has suffered severe degradation. It is likely M. honahona became extinct in the late 1990s, not long after it was first discovered. Despite the similar spelling of the type localities for M. madagascariensis and M. honahona, it is clear from the description and accompanying notes made by Arnoult (1963) that the specimens he collected were from forested swamps located to the north of Toamasina, near Mahambo, which is approximately 650 km (400 miles) to the north of the Réserve Spéciale de Manombo, type locality of M. honahona (fig. 1).
Local Name: None known.
Etymology: Named for the swampy Pandanas dominated habitat in which this species occurred within the Réserve Spéciale de Manombo, in southeastern coastal Madagascar. Hohahona translates as swamp or swampy in Malagasy. The epithet, honahona, is used as a noun in apposition.
FIGURE 2.
Malagodon honahona, new species, holotype, UMMZ 240245, adult male, 23.9 mm SL. Réserve Spéciale de Manombo, southeastern coastal Madagascar. Preserved in ethanol.

FIGURE 3.
Malagodon honahona, new species, allotype, UMMZ 254162, adult female, 18.9 mm SL. Réserve Spéciale de Manombo, southeastern coastal Madagascar. Preserved in ethanol.

TABLE 1.
Morphometric and meristic data for holotype, allotype, and paratypes of Malagodon honahona. M = male; F = female.

FIGURE 4.
Live adult male Malagodon honahona, UMMZ 240245, holotype, 23.9 mm SL, photographed shortly after capture. (Photo by P. Reinthal and J.S. Sparks.)

FIGURE 5.
Comparison of adult male and female cleared-and-stained Malagodon species. Malagodon honahona: A, male, AMNH 278989, 20.2 mm SL, and B, female, AMNH 278989, 18.6 mm SL. Malagodon madagascariensis: C, male, AMNH 20526, 22.3 mm SL, and D, female, MNHN 1963-166, 22.2 mm SL. Asterisks (*) mark the neural spine on the fifth vertebral centrum for comparison. Inset on C is of same specimen with additional alizarin staining to visualize neural spines more easily. Scale bars = 1 mm.

DISCUSSION AND COMPARISONS
As previously noted, the two valid species of Malagodon are easily distinguished from all Recent mainland African species assigned to Pantanodontidae sensu Meinema and Huber (2023), by: (1) the presence of oral-jaw teeth in both sexes (fig. 5); (2) pelvic fins in males that are highly modified, with up to the first three pelvic-fin rays becoming thickened spinelike processes in larger, sexually mature individuals (fig. 6); (3) the presence of hooks, which are frequently paired, on pelvic rays 1–4 in males (hooks continue to develop on additional rays as the individual matures; see developmental progression in fig. 6); (4) an increased pleural-rib count (12–14); (5) the hypural plates in the caudal skeleton are fully fused (fig. 5); and (6) the dorsal and anal fins of males are significantly elongated and produced, with the development of long trailing margins in larger individuals (figs. 2, 4, 7A). Members of Malagodon and Pantanodon also exhibit clear sexual dimorphism in the length and size of their anal fin, with males possessing longer fins with more produced trailing margins. The only other species of fish found in Madagascar that could potentially be confused with Malagodon is an introduced species of Gambusia (G. holbrooki), commonly known as the eastern mosquitofish. However, in the latter taxon, the anal fin of males is modified into a gonopodium, the dorsal and anal fins are not produced, and the pelvic fins of males lack hooks at the tips regardless of size or sexual maturity. Female G. holbrooki are considerably deeper bodied than female members of Malagodon, especially on the anterior half of the body.
FIGURE 6.
Ontogenetic series comparing pelvic-fin development during ontogeny in Malagodon males. Malagodon honahona: A, UMMZ 254159, 20.2 mm SL; C, UMMZ 254161, 18.3 mm SL; E, UMMZ 240245, holotype, 23.9 mm SL. Malagodon madagascariensis: B, MNHN 1963-166, 11.8 mm SL; D, MNHN 1963-166, 21.6 mm SL; F, MNHN 1963-166, 26.1 mm SL. S = spinelike process with associated hook. Numerals 1–4 designate hooked segmented rays in series, following thickened spinelike processes, if present. Both spinelike processes and associated hooks increase in number over ontogeny, with sexually mature males possessing both more spinelike processes and hooks: (B) no thickened spinelike process, no hook; (A) no spinelike process, first ray with single hook; (C, D) one spinelike process present, exhibiting single hook, and following ray with paired hooks; (E) two spinelike processes present, with first spinelike process exhibiting single hook, second with paired hooks, and following ray with paired hooks; and (F) three spinelike processes present, all exhibiting paired hooks, and following ray also with paired hooks.

FIGURE 7.
Malagodon madagascariensis: A, adult male, MNHN 1963-166, paratype, 20.8 mm SL. B, adult female, MNHN 1963-166, paratype, 17.6 mm SL. Forest streams near Mahambo, northeastern coastal Madagascar. Preserved in ethanol.

In addition to the apomorphic features listed above in the Diagnosis for M. honahona, including a lower anal-fin ray count (15–17 vs. 18–19), a longer caudal peduncle (26.8–39.8 vs. 21.9–26.7% SL), and a notably expanded and platelike neural spine on the arch of the fifth vertebral centrum in both sexes (vs. thin and spinelike in M. madagascariensis) (fig. 5A, B), the new species possesses neural spines on both the sixth and seventh vertebral centra that are also somewhat expanded and platelike dorsally (but much less so than the neural spine on the fifth vertebral centra) compared to those in M. madagascariensis, which are notably narrow and spinelike (fig. 5C, D).
The means of the following features in M. honahona, new species, were also determined to be significantly different from M. madagascariensis, despite all exhibiting overlapping ranges (see table 1): caudal-fin rays [25–27 vs. 27, 29 (in a single specimen)], lateral-line scales (28–29 vs. 29–30), caudal-peduncle depth (10.8–15.0 vs. 12.8–16.7% SL), pectoral-fin length (12.0–22.9 vs. 16.5–20.7% SL), dorsal-fin base length (7.5–13.5 vs. 10.9–15.3% SL), pelvic-fin length (8.4–14.2 vs. 7.9–13.6% SL), anal-fin base length (16.5–24.7 vs. 20.5–27.4% SL), and snout length (17.7–30.4 vs. 15.0–27.8% HL) (figs. 2–5, 7).
Despite repeated attempts over the past three decades, no additional specimens of M. honahona have been collected from the type of locality or other potentially suitable habitat in the region, and therefore the species is presumed to be extinct. The Réserve Spéciale de Manombo preserved some of the last remaining coastal rainforest in southeastern Madagascar. However, despite this protection, the reserve and surrounding region, including the upstream watershed that fed the small swamp at the type locality, have suffered severe deforestation and environmental degradation since the late 1990s. This, in combination with competition from exotic species such as G. holbrooki, which were far more common at the type locality than the new species even in 1994, likely contributed to the rapid extinction of M. honahona not long after the species was first discovered.
ACKNOWLEDGMENTS
We are especially grateful to K. Riseng for considerable assistance with fieldwork and to P. Bragança (AMNH) and M. Ghedotti (Regis University) for generously sharing their knowledge of cyprinodontiform anatomy. Particular thanks to P. Reinthal for recognizing the new species as a member of Pantanodon at the time of collection. We thank D. Nelson and R. Singer (UMMZ), G. Duhamel, P. Pruvost, and R. Causse (MNHN), and D. Catania (CAS) for the loan or gift of specimens in their care. The comments of K. Tang (University of Michigan) and an anonymous reviewer greatly improved this manuscript. Fieldwork in Madagascar was facilitated by B. Andriamahaja and the MICET (Institute for the Conservation of Tropical Environments, Madagascar) staff, and we are grateful for their continued support of ichthyological field research in Madagascar. Collection permits were obtained from the Direction des Eaux et Forêts and the Association National pour la Gestion des Aires Protégées (ANGAP), Madagascar.
Copyright © American Museum of Natural History 2024
REFERENCES
Appendices
APPENDIX 1
Material Examined
Number of specimens following catalog numbers indicate count of individuals examined in this study and do not necessarily correspond to the total number of specimens in any particular lot.
Malagodon madagascariensis: AMNH 20526: Paratypes; 3 ex. (1 ex. C&S male), 17.6–22.4 mm SL; Madagascar: Toamasina Province: East Coast: north of Toamasina (Tamatave): forest streams of Mahambo; J. Arnoult, X-1962. MNHN 1963-166: Paratypes; 5 ex. (males), 15.9–27.4 mm SL; Madagascar: Toamasina Province: East Coast: north of Toamasina (Tamatave): forest streams of Mahambo; J. Arnoult, X-1962. MNHN 1963-166: Paratypes; 5 ex. (females), 14.5–22.9 mm SL (1 ex. C&S); Madagascar: Toamasina Province: East Coast: north of Toamasina (Tamatave): forest streams of Mahambo; J. Arnoult, X-1962.
Pantanodon podoxys: CAS (SU) 50194: Holotype; 1 ex., male; 35.1 mm SL; Africa: Tanzania: Pwani Region: in swampy land a few miles inland from Dar-es-Salaam; A. W. Herre. CAS (SU) 61761: 1 ex., 31.6 mm SL; Africa: Tanzania: Dar-es-Salaam Region: Msimbazi Creek; A. W. Herre, 26 Apr. 1934.
Pantanodon propinquus: CAS (SU) 62115; 2 ex., 16.2–30.1 mm SL; Africa: Tanzania: Tanga Region: 20 miles north of Tanga; J. H. E. Leakey, 23 Aug. 1963.